
Adibi Laboratory
Dr. Adibi's research consists of biomedical studies, including devices, to solve diagnostic challenges and provide treatment options for Temporomandibular Disorders and Nocturnal Bruxism (nighttime grinding of teeth).

Explore the dynamic research labs at UTHealth Houston School of Dentistry, where faculty lead innovative studies across the oral, craniofacial, and dental sciences. These labs span a broad spectrum of inquiry — from basic and translational science to clinical and community-based research — uniting faculty, students, and collaborators in advancing the biological, behavioral, and technological foundations of oral health.
Dr. Adibi's research consists of biomedical studies, including devices, to solve diagnostic challenges and provide treatment options for Temporomandibular Disorders and Nocturnal Bruxism (nighttime grinding of teeth).
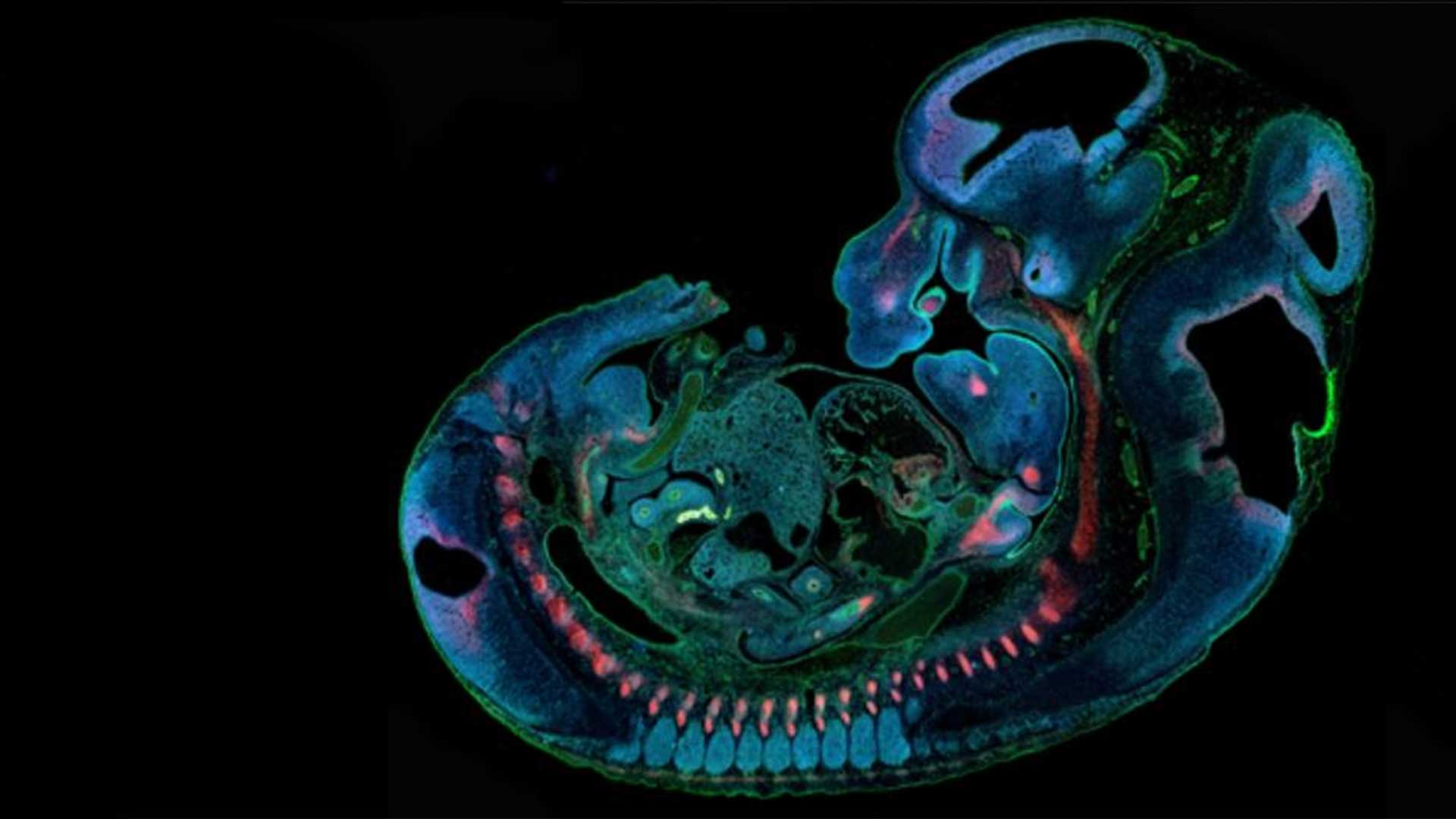
Dr. Fakhouri’s research investigates the genetic and epigenetic factors essential to early craniofacial development and disruptions in these factors that cause and contribute to congenital craniofacial defects, including cleft lip and palate.
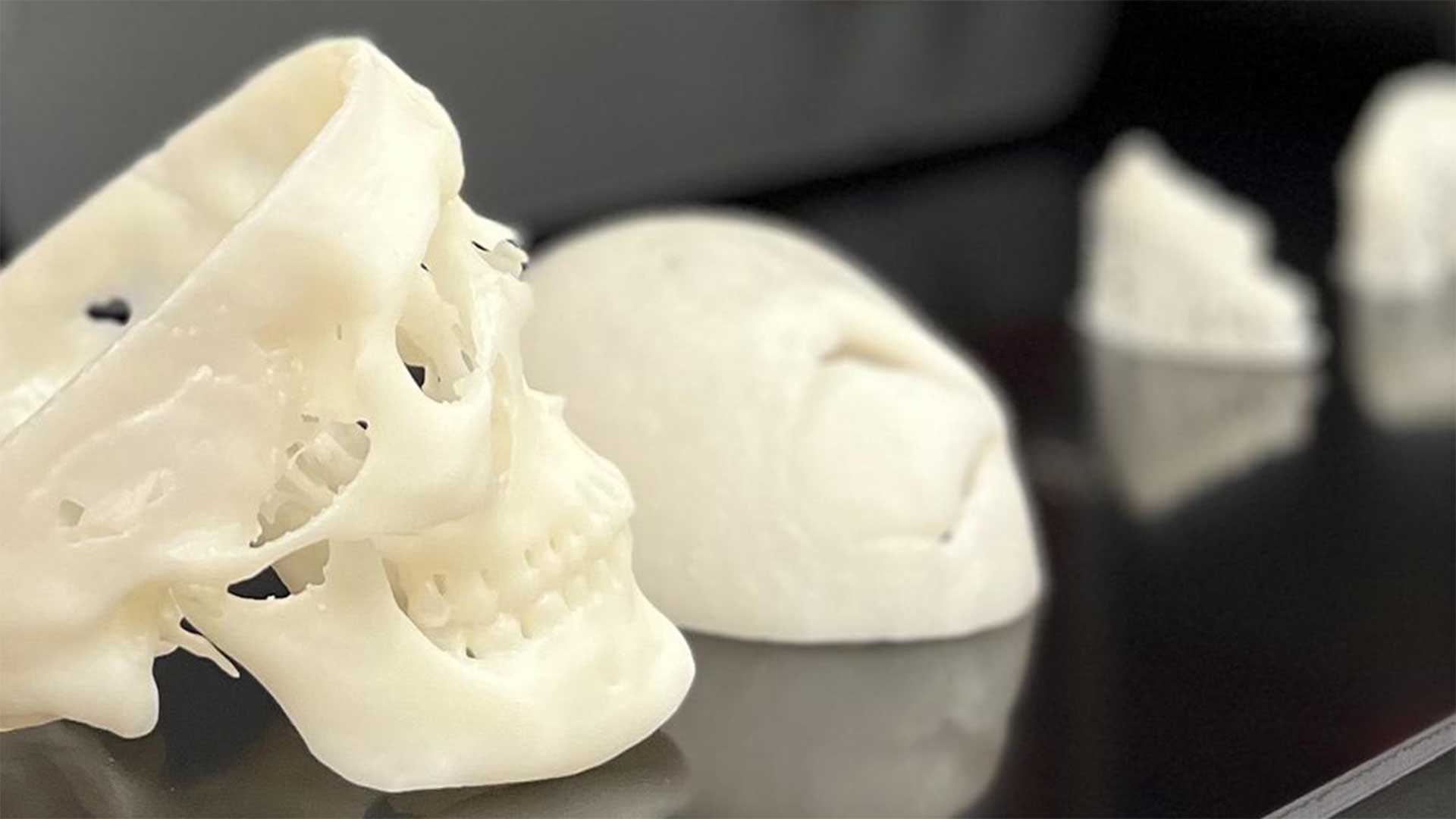
Dr. Kasper applies fundamentals of engineering, materials science, and the biosciences to the development and evaluation of biomaterials-based technologies to meet a variety of clinical needs, with a particular focus on 3D printing and tissue regeneration.
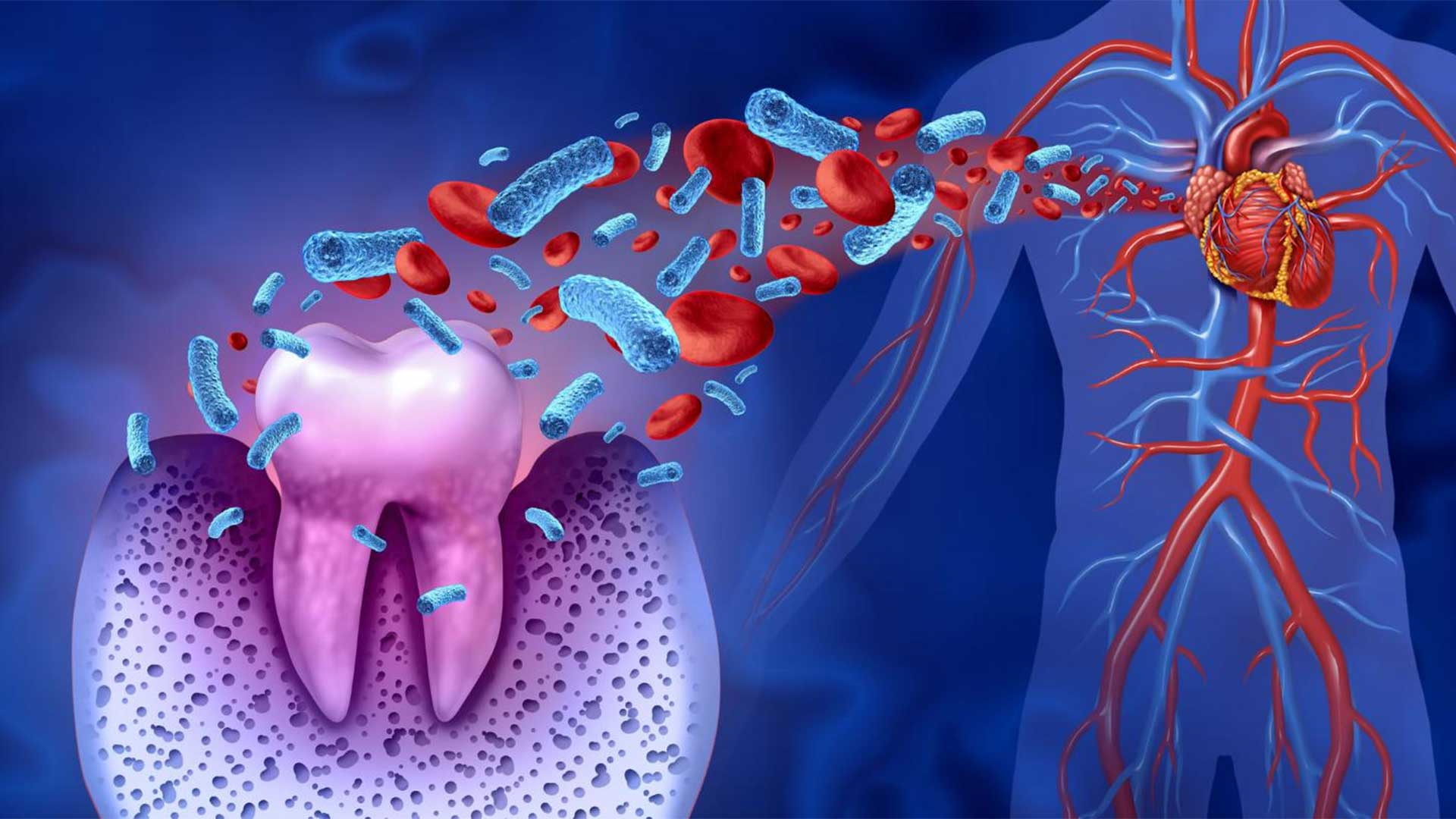
Dr. Lee’s research focuses on understanding the resolution of inflammation in periodontitis and on investigating novel approaches to diagnose and treat periodontal diseases.
Dr. Myers’s research tackles adverse health consequences of toxicokinetic drug interactions with drugs, medicinal therapies, and drugs of abuse; as well as increasing the body of knowledge about the metabolism/excretion of drugs to assist smoking cessation.
Drs. Wanida and Noriaki Ono’s research in the Tooth Lab use advanced genetically engineered mouse model approaches to address fundamental questions on the growth, development, health, and diseases of bones and teeth.
Dr. Young’s research seeks to develop material-based immunotherapies for the treatment of head and neck cancer and explores the mechanisms of how immunotherapy may synergize with other modalities of treatment such as chemo/radiation therapy to improve overall survival and quality of life.